
L’idée qu’à une séquence en acides aminés correspond un unique repliement protéique (dite aussi principe d’Anfinsen, du nom du biochimie et lauréat du prix de Nobel de Chimie 1972 Christian Anfinsen) a longtemps joué un rôle central en biologie structurale. Et puis au fil des décennies, de plus en plus d’exceptions à cette règle ont fait leur apparition, qu’il s’agisse de protéines désordonnées, ou bien de protéines capable de changer de forme, pour le meilleur (la ranaspumine des nids de grenouilles) comme pour le pire (les terribles amyloïdes).
En opposition aux protéines monomorphes (qui n’adoptent qu’un unique repliement), il existe également des protéines métamorphes, dont la structure peut alterner de manière réversible entre deux repliements distincts. Le métamorphisme doit donc être distingué de l’allostérie, où la protéine va changer de forme de manière limitée et en conservant ses éléments de structure secondaire, et de la formation de fibres amyloïdes, qui est un phénomène irréversible. Si les protéines métamorphes sont connues depuis le début des années 80, elles restent néanmoins rarissimes au sein de la PDB (avec environ une petite centaine d’exemples connus), les chercheur·se·s estiment que 0.5 à 4% des protéines seraient dotées de ce super pouvoir, mais l’apparition de celui-ci au cours de l’évolution reste encore mal comprise.

La question que tout le monde se pose : Métamorph est il entièrement constitué de protéines métamorphiques ?
L’un des exemples les plus frappant des ces métamorphes est la lymphotactine, dite aussi chemokine XCL1, une protéine de la famille des cytokines qui est impliquée dans la migration et le positionnement des cellules du système immunitaire. Celle-ci peut en effet être présente dans la cellule sous la forme d’un monomère avec un repliement mixte α+β typique des chémokines, ou sous forme de dimère avec uniquement des feuillets β. La prédominance d’une forme ou l’autre dépendra alors de l’environnement cellulaire. Ainsi, une faible concentration en sel et des hautes températures favorisent-elles la formation du dimère par rapport au monomère.
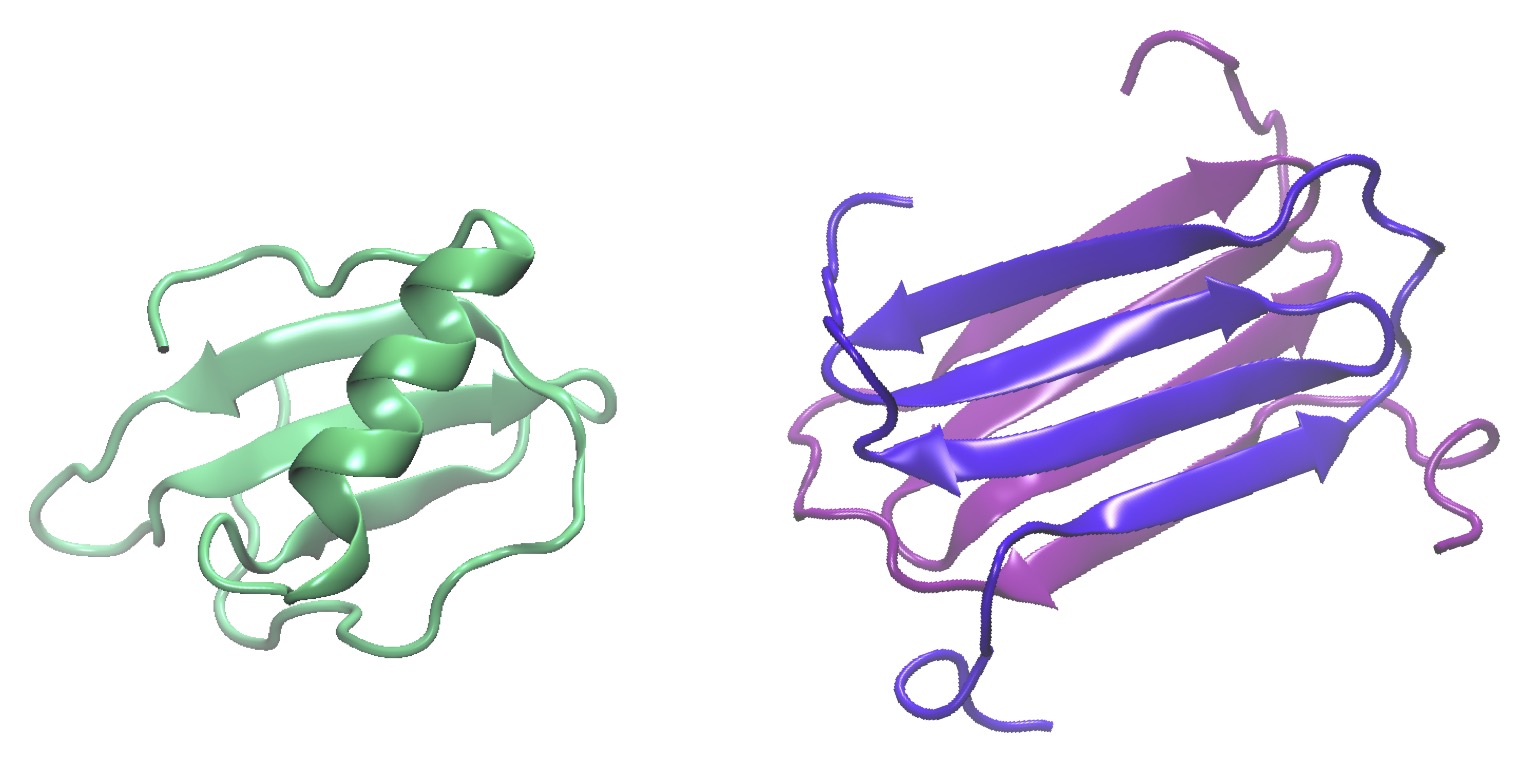
Les deux structures adoptées par la lymphotactine, soit sous forme de monomère (en vert, code pdb 4hed) ou de dimère (en violet, pdb 2jp1)
Très récemment, des équipes de recherche ont entrepris de reconstituer l’arbre généalogique de la lymphotactine pour remonter aux sources de son métamorphisme. Pour ce faire, elles ont étudié les séquences de plus de 450 chémokines apparentées à XCL1, jusqu’à pouvoir reconstituer les séquences de plusieurs ancêtres de notre métamorphe. L’écrasante majorité des chémokines sont monomorphes, notamment grace à deux ponts disulfure qui verrouillent la structure dans sa forme α+β et empêchent un changement de repliement. Et c’est également le cas des grands ancêtres de XCL1, mais en descendant l’arbre on trouve des ancêtres, que l’on nommera pépé Prot1 et pépé Prot2, qui ont perdu l’un de ces ponts et gagné la capacité d’adopter un repliement alternatif. Ces aieuls ne possèdent néanmoins pas la même souplesse que leur descendance quand il s’agit de se transformer. Ainsi pépé Prot1 préfère-t-il nettement le repliement de type chémokine, tandis que pépé Prot2 a un faible prononcé pour la formation du dimère. XCL1 quant a elle est capable de naviguer avec aisance d’une forme à l’autre. Mais au fait, pour quoi faire ?

Au risque de décevoir le lectorat de ce blog (et de briser des vocations), je me dois de vous informer que l’obtention d’une séquence ancestrale de protéine ne nécessite pas de brandir un glaive en prononçant une formule magique (et c’est bien dommage)
Il semble que le métamorphisme protéique puisse présenter plusieurs intérêts évolutifs. Il permet déjà une économie de moyens, puisque qu’un gène unique va coder une protéine capable de remplir différentes fonctions selon la forme qu’elle adoptera. De plus, le changement de repliement permet à la cellule de moduler l’activité de ses protéines plus rapidement (en quelques secondes) que s’il fallait produire deux protéines distinctes. Ainsi, dans le cas de la lymphotactine, celle-ci peut soit jouer son rôle de cytokine lorsqu’elle est sous sa forme monomère, soit avoir une activité antimicrobiale quand elle s’assemble en dimère. Un double effet Kiss Cool précieux quand il s’agit de lutter contre des pathogènes ! On comprend donc que les métamorphes attirent l’attention des chercheur·se·s, puisqu’elles sont une source d’inspiration pour le développement de nouvelles molécules thérapeutiques capables d’agir à plusieurs niveaux.
Une erreur d’étymologie ?
Dans la mythologie grecque, Protée/Proteus était une divinité marine dotée du pouvoir de métamorphose. C’est notamment à lui que l’on doit l’adjectif protéiforme. Mais par contre il n’a rien à faire avec nos protéines, qui elles tiennent leur nom d’un autre mot grec prôtos, signifiant premier. Alors, se pourrait-il que les protéines métamorphes se soient emmêlées les pinceaux dans l’origine de leur nom ?
