
Après l’odorat, le goût, la vue et le toucher, voici donc notre ultime billet dédié aux protéines qui sont associées à nos cinq sens, et cette fois nous allons parler audition !
Au sein de la cochlée, ce petit os en forme de coquille d’escargot logé au coeur de l’oreille interne des mammifères, on trouve des cellules ciliées externes, dont la fonction est d’amplifier le signal sonore entrant en se déformant. Et dans ces cellules ciliées externes, on trouve la prestine, une protéine motrice qui va être impliquée dans la réponse cellulaire aux ondes sonores. Elle contient deux domaines, un canal échangeur d’ions ancré dans la membrane cellulaire, qui va permettre à des ions chlorures (Cl-) d’entrer dans la cellule tandis que des carbonates (CO32-) vont quitter celle ci, et un domaine cystoplasmique, où un chlorure pourra se fixer. Cette fixation du chlorure entraine une déformation de la protéine, qui à son tour va induire un changement de forme de la cellule ciliée. Contrairement aux autres protéines motrices, comme la kinésine ou la dynéine, dont les mouvements sont dus à l’énergie fournie par une réaction chimique (l’hydrolyse de l’ATP), le mouvement de la prestine provient de la fixation d’un ion, ce qui en fait un capteur du potentiel électrique de la cellule.
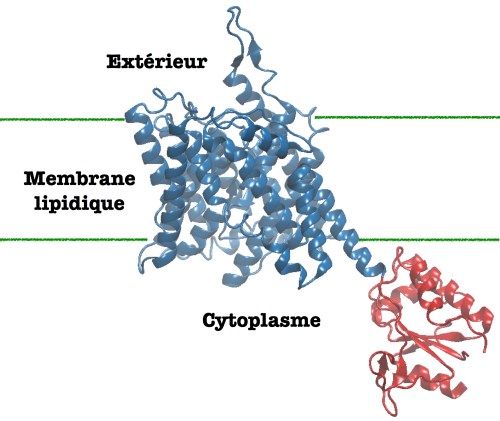
Découverte il y a une vingtaine d’années par une équipe de recherche américaine, la prestine doit son nom au terme musicale Presto, qui permet de souligner sa rapidité d’action, qui est de quelques µs, alors que ses collègues protéines motrices nécessitent plutôt quelques ms pour fonctionner. L’absence de cette protéine ou des mutations de celle-ci chez un individus seront alors associées à une perte des ses capacités auditives. Les ions salicylates (qui sont des dérivés de l’acide acetylsalicylique, soit l’aspirine) sont également susceptibles de se fixer à la prestine, ce qui peut expliquer les troubles auditifs observés chez des patients en cas d’intoxication à l’aspirine.

La prestine joue donc un rôle central pour l’audition des mammifères, mais il semble qu’elle soit également impliquée dans les capacités d’écholocation de certaines espèces animales.
L’écholocation, c’est cette stratégie qui consiste à envoyer des sons, le plus souvent des ultra-sons avec des fréquences supérieures à 20kHz, et analyser la façon dont ceux-ci reviennent vers l’émetteur pour percevoir son environnement. C’est la technique à l’origine du sonar des sous-marins, et cette faculté est particulièrement développée chez les chauve-souris et les cétacées. Il y a une dizaine d’années, des chercheur·se·s ont comparé la séquence de la prestine chez plus d’une cinquantaine d’espèces de mammifères (capables ou non d’écholocation) et ont montré que les mutations observées entre les séquence sont directement liées à la capacité de percevoir les hautes fréquences chez les espèces considérées. Qui plus est, on observe des modifications identiques de la séquence protéique au cours de l’évolution pour les chauve-souris et les baleines. On a donc un beau cas d’évolution convergente, où des familles animales distinctes possèdent une protéine qui a évolué de manière similaire pour exercer la même fonction (soit ici la perception des hautes fréquences sonores qui permet l’écholocation).
