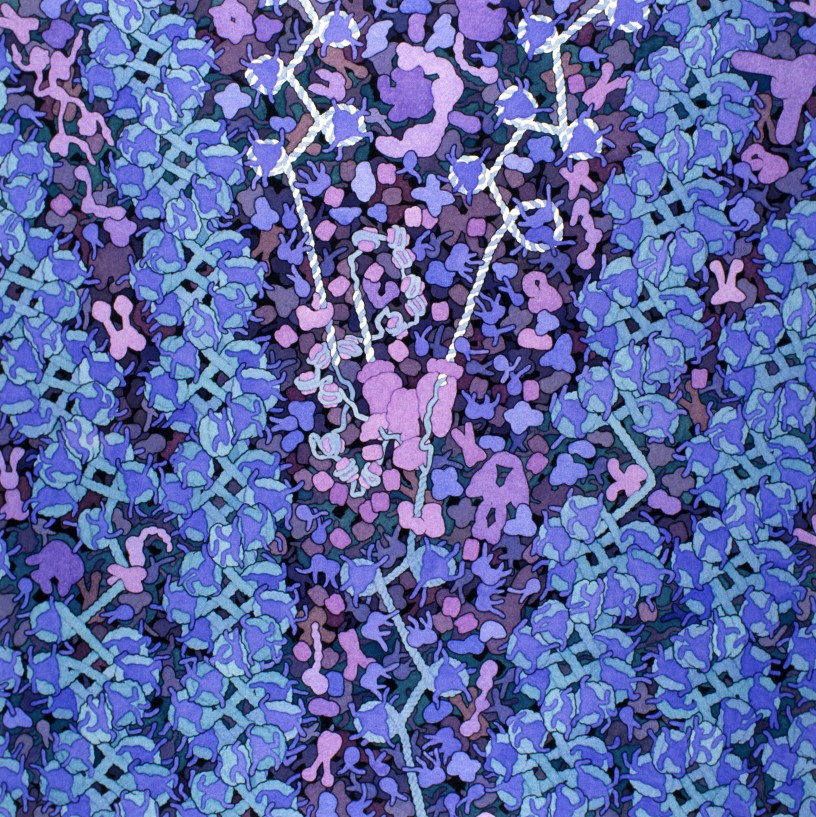
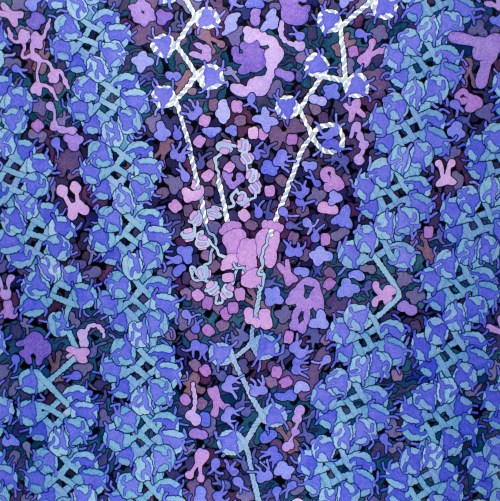
Dans un des tous premiers billets de ce blog, nous avions comparé l’ADN de nos cellules à une grande bibliothèque du vivant, qui comprendrait toutes les instructions nécessaires à la fabrication et l’entretien d’un organisme. Bien entendu, cette bibliothèque il faut que quelqu’un veille à ce qu’elle soit correctement rangée.
Dans les cellules humaines par exemple, si l’on déroulait le filament d’ADN qui constitue nos 23 paires de chromosomes, on obtiendrait un long brin comprenant plus de 3 milliards de paires de base, de seulement 2 nm (soit 10-9 m !) de diamètre, mais qui mesure près de 2 m de long. Ce qui semble un peu difficile à caser dans un noyau cellulaire qui fait lui même environ 10 μm de diamètre !
Et pourtant, une fois l’ADN correctement embobiné sous forme de chromosomes, ceux-ci ne font plus que quelques μm de long, soit un taux de compactage de l’ordre du million. Cet exploit d’organisation et de rangement à faire pâlir une grande enseigne de meubles suédoise, c’est à la chromatine qu’on le doit.

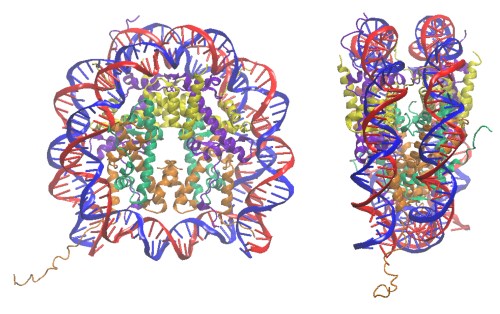
La chromatine, c’est donc un complexe formé par l’association de l’ADN avec un certain nombre de protéines (dont notamment les histones), et qui va permettre la compaction de celui-ci dans le noyau des cellules eucaryotes (les cellules procaryotes, qui sont dépourvues de noyau, ont adopté un autre système de rangement de l’ADN que nous n’aborderons pas dans ce billet). L’empaquetage se fait par plusieurs niveaux d’enroulement successifs, en démarrant par un double tour d’ADN autour des histones, ce qui va former le nucléosome. Au coeur du nucléosome on trouve huit molécules d’histones (de quatre variétés présentes en deux exemplaires chacune). Ces protéines sont riches en acides aminés portant des charges positives (tels que la lysine ou l’arginine), et qui vont fortement s’associer avec les charges négatives portées par la molécule d’ADN.
Après quoi on a encore plusieurs niveaux d’embobinage, qui sont maintenus en place via des interactions entre l’ADN et différentes protéines, jusqu’à aboutir au chromosome. Cette façon de ranger l’ADN produit un assemblage très dense, et cette densité va déterminer l’accessibilité du matériel génétique pour sa transcription en ARN. On distingue donc souvent deux types de chromatines, l’hétérochromatine, très compacte, où l’ADN est difficile d’accès, et l’euchromatine, moins compacte et où l’ADN polymérase peut accéder aux gènes. La densité de la chromatine peut donc varier localement et au cours du temps, en fonction de petites modifications au niveau des histones, ce qui permet de moduler l’expression (i.e. la transcription, puis la traduction en protéines) des gène selon les besoins de la cellule à un moment donné. L’empaquetage de l’ADN dans la chromatine a également un rôle protecteur pour celui-ci (même si la cellule est équipée pour le réparer et poser des rustines si nécessaire, elle préfère néanmoins que son ADN reste intact).
La chromatine a été découverte à la fin du dix-neuvième siècle, et doit son nom à sa capacité à fixer les colorants (du grec chroma pour la couleur). Les histones furent quant à eux découverts en 1884 par Albrecht Kossel. Mais il a fallu attendre les années 1960 pour que l’on commence à pouvoir observer les structures de ces assemblages par microscopie électronique, ce qui a valu un Prix Nobel de Chimie à Aaron Klug en 1982. Enfin, la structure du nucléosome a été résolue par cristallographie en 1997 par Karolin Luger.
Au final la chromatine a donc toutes les qualités d’une bibliothécaire de choc. Elle est en charge du bon rangement et de l’intégrité du matériel génétique de la cellule, tout en devant également s’assurer que celui-ci reste accessible quand le besoin s’en fait sentir.
Il ne reste plus qu’à savoir si les protéines qui pénètrent dans le noyau sont tenues de parler à voix basse !

{kind=link}